Speciacija: nastanek vrst v evoluciji, mehanizmi in primeri
Speciacija: poglobljen vodič o nastanku vrst v evoluciji — mehanizmi (alopatrija, simpatrija, hibridizacija), primeri in DNK dokazi za razumevanje nastanka vrst.
Speciacija se nanaša na to, kako se vrste oblikujejo in ločijo v naravi. Je osnovni proces v evolucijski biologiji, saj pojasnjuje nastanek biotske raznovrstnosti in razhajanja vrstnih linij skozi čas.
Galerija slik
10 Slike









Zgodovinski pogled: anageneza in kladogeneza
Že Darwin je razmišljal o načinu nastajanja vrst. Opisal je dve pogosti ideji:
- Anageneza – spreminjanje ali "fitoletična evolucija" znotraj ene razvojne linije, pri kateri ena vrsta postopoma prehaja v drugo brez delitve v več sodobnih vrst.
- Kladogeneza – razvejanje ali delitev enega prednika na dve ali več ločenih vrstnih linij; danes velja kladogeneza za glavni mehanizem nastanka novih vrst.
Glavni mehanizmi speciacije
Speciacija je pogosto povezana z mehanizmi izolacije, ki omejijo genski pretok med populacijami. Med najpomembnejšimi so:
- Geografska izolacija (alopatrija) – fizična ločitev populacij (gore, reke, oceani, podnebne spremembe), kar omogoči neodvisno evolucijo in sčasoma reproduktivno izolacijo.
- Parapatrija – sosednje populacije ob meji z okolji, kjer se selekcijski pritiski razlikujejo; genski pretok je možen, a selekcija vodi k lokalni prilagoditvi in delitvi.
- Sympatrija – nastanek novih vrst znotraj istega geografskega območja, pogosto zaradi močne ekološke diferencijacije (npr. različen izbor hrane) ali sprememb v reproduktivnem vedenju.
- Peripatrija – posebna oblika alopatrije, kjer majhna odcepljena populacija na robu razširjenega območja hitro diverzira zaradi genetike majhne populacije in močne selekcije.
Reproduktivna izolacija: pred in po oploditvi
Reproduktivna izolacija vključuje skupek lastnosti, ki preprečujejo medsebojno razmnoževanje ali zmanjšujejo plodnost potomcev. Delimo jo na:
- Prezygotne (pred oploditvijo): časovna/ekološka ločitev, vedenjska imkopatibilnost (drugačna pheromonska ali gibalna simfonija), mehanske ovire, gametne nezdružljivosti.
- Postzygotne (po oploditvi): zmanjšana preživetnost hibridov, sterilnost (npr. mule), ali zmanjšana fitnes naslednjih generacij.
Proces, ko se zamenjanimi izbiri ojača reproduktivna izolacija, se imenuje reinforcement.
Vloga hibridizacije in genske izmenjave
V zadnjih desetletjih so analize zaporedja DNK pokazale, da med sorodnimi vrstami pogosto prihaja do križanja. Preko takih stikov se lahko prenašajo geni. To pomeni, da reproduktivna izolacija ni vedno absolutna in da nastanek vrste ni nujno vedno posledica popolne geografske ločitve (alopatrije). Nekateri pomembni pojavi so:
- Introgression – vnos posameznih genov iz ene vrste v drugo skozi ponavljajoča se križanja in zadnja križanja nazaj v populacijo.
- Hibridna speciacija – nastanek nove vrste iz križanja dveh ločenih vrst; pogosta pri rastlinah (poliploidija), včasih pa tudi pri živalih (homoploidna hibridna speciacija).
- Genomska mozaicnost – sodobne genomike razkrivajo, da so številne vrste genomskimi "mozaiki" več izvornih linij, kar vpliva na fenotip in ekologijo.
Ekološka in spolna selekcija kot gonilna sila
Speciacija pogosto poteka kot posledica ekološke diferencijacije (ekološka speciacija), kjer različne okoljske razmere izbirajo za različne lastnosti. Spolna selekcija (npr. izbor partnerjev na podlagi barve, pesmi ali vedenja) lahko dodatno potisne populacije k hitrejši reproduktivni izolaciji.
Primeri iz narave
- Darwinove naše (finches) na Galapagosu – kladogeneza, pricem so diferenciacije v kljunih in vedenju pomenile hitro prilagoditev različnim virom hrane.
- Afriška kupljena jezera (cichlidi) – eksplozija vrstne raznovrstnosti preko ekološke speciacije in spolne selekcije.
- Muha Rhagoletis pomonella – primer sympatrične speciacije, kjer so populacije, ki se hranijo na jabolkih namesto na hruškah, začele divergirati.
- Rastline z akutno poliploidijo (npr. nekatere vrste iz rodu Tragopogon ali pšenica) – nova poliploidna vrsta se lahko pojavi v eni generaciji in je takoj reproduktivno izolirana od starševskih vrst.
- Ring species (npr. nekatere vrste galebov ali salamanders iz rodu Ensatina) – primer postopnega divergiranja vzdolž geografske krožnice, kjer se sosednje populacije križajo, medtem ko se skrajni klini ne križata.
Kaj so molekularne študije pokazale?
Genomske in filogenetske analize so razkrile, da je speciacija pogosto zapleten in mozaik proces: nekatere genske regije divergirajo hitro pod močnim selekcijskim pritiskom, medtem ko druge ostajajo združljive in kažejo znake genske izmenjave. Pojavi, kot sta nepopolno združevanje linij (incomplete lineage sorting) in hibridizacija, otežujejo rekonstrukcijo čiste, vejatvne zgodovine, vendar hkrati bogatijo genski sklad vrst.
Zaključek
Speciacija ni enotna ali vedno linearna: vključuje kombinacijo geografskih, ekoloških, gensko-driftnih in vedenjskih procesov. Medtem ko je fizična ločitev pogosto močan in pogost dejavnik, sodobne molekularne študije kažejo, da so hybridizacija, introgressija in selektiven pretok genov prav tako pomembni. Celostno razumevanje speciacije zahteva kombinacijo poljskih opazovanj, eksperimentalnih študij in genomske analize, ki skupaj pojasnijo, kako nastajajo nove vrste in kako se ohranja biotska raznovrstnost.
Izolacijski mehanizmi
Izolacijski mehanizmi so stvari, ki preprečujejo uspešno razmnoževanje med skupinami v vrsti. Vzpostavi se reprodukcijska izolacija populacij. To je še posebej pomembno za koncept bioloških vrst, saj so vrste opredeljene z razmnoževalno izolacijo.
Izolacijske mehanizme lahko razdelimo v dve skupini, pred oploditvijo in po njej.
Pred oploditvijo
Dejavniki, ki posameznikom preprečujejo parjenje.
- Geografska izolacija: Vrste se pojavljajo na različnih območjih in so pogosto ločene z ovirami.
- Časovna izolacija: Posamezniki se ne parijo, ker so aktivni ob različnih časih. To so lahko različni deli dneva ali različni letni časi. Obdobja parjenja med vrstami se ne ujemajo. Posamezniki se ne srečujejo med seboj v obdobju parjenja ali sploh ne.
- Ekološka izolacija: Posamezniki se parijo le v svojem najljubšem habitatu. Ne srečujejo osebkov drugih vrst z drugačnimi ekološkimi preferencami.
- Vedenjska izolacija: Posamezniki različnih vrst se lahko srečajo, vendar ne prepoznajo morebitnih spolnih znakov. V večini primerov posameznik izbere pripadnika svoje vrste.
- Mehanska izolacija: Pri tem lahko pride do poskusa kopulacije, vendar do prenosa sperme ne pride. Osebki so lahko nezdružljivi zaradi velikosti ali morfologije.
- Igralna nezdružljivost: Pri tem pride do prenosa sperme, vendar jajčece ni oplojeno.
Po oploditvi
Dejavniki, ki preprečujejo uspešno parjenje, kot so genetska nezdružljivost, hibridna nesposobnost ali sterilnost.
- Zygotična umrljivost: Jajčece je oplojeno, vendar se zigota ne razvije.
- Neživljenjskost hibrida: hibridni zarodek se oblikuje, vendar ni sposoben za življenje.
- Hibridna sterilnost: hibrid je sposoben življenja, vendar je odrasli osebek sterilen.
- Hibridna razčlenitev: Prva generacija hibridov (F1) je vitalna in plodna, nadaljnje generacije hibridov (F2 in povratni križanci) pa so neživljenjske ali neplodne.

Geografska izolacija
To naj bi bil najpogostejši vzrok za nastanek vrste. Prvi je na to pomislil Moritz Wagner, nemški raziskovalec in naravoslovec.
Wagner je na začetku svoje kariere deloval kot geograf in objavil več geografskih knjig o Severni Afriki, Bližnjem vzhodu in Tropski Ameriki. Bil je tudi navdušen naravoslovec in zbiratelj, po tem delu pa je med biologi najbolj znan. Ernst Mayr, evolucionist in zgodovinar biologije, je opisal Wagnerjev pomen. str. 562-565.
Med triletnim bivanjem v Alžiriji je Wagner med drugim preučeval neleteče hrošče Pimelia in Melasoma. Vsak rod je razdeljen na več vrst, od katerih je vsaka omejena na del severne obale med rekami, ki se z gorovja Atlas spuščajo v Sredozemsko morje. Takoj ko prestopimo reko, se pojavi druga, a tesno sorodna vrsta.
"... začetna vrsta se bo [pojavila] le, ko bo nekaj posameznikov prestopilo mejne meje svojega areala... oblikovanje nove rase ne bo nikoli uspelo... brez dolgotrajne ločitve kolonistov od drugih pripadnikov njihove vrste".
To je bil zgodnji opis procesa ene vrste geografske speciacije. Leta 1942 ga je ponovno uvedel Mayr in pomen geografske speciacije je postal ena od osrednjih idej evolucijske sinteze.
Drugi izraz za geografsko speciacijo je alopatrična speciacija. Allopatrija pomeni "različna dežela".
Prosti otoki
Vulkanski otoki so nastali brez življenja, vse življenje pa mora priti z vetrom ali vodo. Na havajskih in galapaških otokih vemo, da se vse oblike življenja spremenijo, ko pridejo na otoke s celine.
Na Havajskih otokih, ki merijo približno 6.500 kvadratnih milj (17.000 km2), živi najrazličnejša zbirka drozofilnih mušic na svetu, od deževnih gozdov do gorskih travnikov. Poznamo približno 800 havajskih vrst drozofilov.
Študije kažejo jasen "pretok" vrst s starejših na novejše otoke. Obstajajo tudi primeri naseljevanja nazaj na starejše otoke in preskakovanja otokov, vendar so ti veliko redkejši.
Z radioaktivnim datiranjem s kalijem/argonom so sedanji otoki nastali pred 0,4 milijona let (Mauna Kea) do 10 milijonov let (Necker). Najstarejši član havajskega arhipelaga, ki je še vedno nad morjem, je atol Kure, ki je datiran na 30 mio let.
Sam arhipelag, ki je nastal zaradi premikanja pacifiške plošče čez vročo točko, obstaja že veliko dlje, vsaj od krede naprej. Havajski otoki in nekdanji otoki, ki so zdaj pod morjem, sestavljajo verigo havajsko- cesarskih podvodnih gora, številne podvodne gore pa so gojzarji.
Vse avtohtone vrste drozofilov na Havajih očitno izvirajo iz ene same predniške vrste, ki je pred približno 20 milijoni let kolonizirala otoke. Kasnejše prilagoditveno sevanje je bilo spodbujeno s pomanjkanjem konkurence in široko paleto prostih niš. Čeprav bi bilo mogoče, da bi otok naselila ena sama breja samica, je bolj verjetno, da je šlo za skupino iz iste vrste.
Na havajskem arhipelagu so tudi druge živali in rastline, ki so doživele podobne, čeprav manj spektakularne prilagoditvene radiacije.
Vrste obročev
V biologiji je obročkasta vrsta povezana vrsta sosednjih populacij, od katerih se lahko vsaka križa s sosednjimi populacijami. Oba konca verige se prekrivata.
Končni populaciji v seriji sta preveč oddaljeni, da bi se lahko križali. Takšne "končne" populacije, ki se ne križajo, čeprav so genetsko povezane, lahko sobivajo na istem območju in tako sklenejo "obroč".
Obročaste vrste so pomemben dokaz evolucije: ponazarjajo, kaj se sčasoma zgodi, ko se populacije genetsko razlikujejo. Richard Dawkins je opazil, da nam obročkaste vrste "le v prostorski dimenziji kažejo nekaj, kar se mora vedno dogajati v časovni dimenziji".
Vendar pa je težko najti preprost in enostaven primer.
Čajke Larus
Klasičen primer obročkaste vrste je obroček cirkumpolarne vrste galebov Larus. Območje razširjenosti teh rac tvori obroč okoli severnega tečaja, ki ga posamezne race običajno ne preletijo.
V splošnem se čuki lahko do neke mere križajo s sosednjimi vrstami, razen na obeh koncih obroča.
Manjše črnoglavke in slede se med seboj dovolj razlikujejo, da se običajno ne križajo, zato skupina čiger tvori kontinuum, razen tam, kjer se obe liniji srečata v Evropi.
Čeprav je resničnost veliko bolj zapletena, dobro prikazuje splošno idejo.
Salamandre Ensatina
Salamandra Ensatina je obročkasta vrsta v gorah okoli kalifornijske Centralne doline. Kompleks tvori podkvasto obliko okoli gora. Čeprav se lahko križajo med 19 populacijami okoli podkve, se podvrsta Ensatina eschscholtzii na zahodnem koncu podkve ne more križati s podvrsto Ensatina klauberi na vzhodnem koncu. To je ilustracija "skoraj vseh stopenj procesa speciacije" (Dobzhansky). Richard Highton je trdil, da gre v primeru Ensatina za več vrst in ne za kontinuum ene vrste.
Zelenkasta pevka
Zelenkasti pevec (Phylloscopus trochiloides) ima več podvrst, od katerih je v Evropi najbolj poznan P. t. viridianus. Gre za obročkasto vrsto, katere populacije se razhajajo vzhodno in zahodno od Tibetanske planote, pozneje pa se srečajo na severni strani. Njihovi odnosi so precej zmedeni.




Simpatrična speciacija
Simpatrična speciacija pomeni nastanek dveh ali več vrst potomcev ene predniške vrste, ki živijo na isti geografski lokaciji. Danes velja, da je to zelo pogost pojav.
Pri simpatrični speciaciji se vrste razlikujejo, medtem ko živijo na istem mestu. Pogosto navedeni primeri simpatrične speciacije so žuželke, ki postanejo odvisne od različnih gostiteljskih rastlin na istem območju.
Obstoj simpatrične speciacije kot mehanizma speciacije je bil zelo sporen. Ljudje so trdili, da so dokazi o simpatrični speciaciji dejansko primeri mikrogeografske speciacije. Na splošno pa to zdaj ni več najprimernejša razlaga. Namesto tega gre za rezultat križanja med tesno sorodnimi vrstami, ki mu sledi naravna selekcija, ki deluje na potomce takšnih križanj. Splošno sprejet primer simpatrične speciacije je primer cichlid iz jezera Nabugabo v vzhodni Afriki, ki naj bi bil posledica spolne selekcije.
Specifikacija s poliploidizacijo
Poliploidija je povzročila številne hitre dogodke speciacije, saj potomci na primer tetraploidnih x diploidnih paritev pogosto dobijo triploidne sterilne potomce.
Vendar pa vsi poliploidi niso reproduktivno izolirani od starševskih rastlin in še vedno lahko pride do pretoka genov, na primer s parjenjem triploidnih hibridov z diploidi, pri katerem nastanejo tetraploidi.
Številne obstoječe rastlinske in večina živalskih vrst so v svoji evolucijski zgodovini očitno doživele poliploidizacijo. Razmnoževanje uspešnih poliploidnih vrst je včasih aseksualno, s partenogenezo. Iz neznanih razlogov je veliko aseksualnih organizmov poliploidnih.
Glogova muha
Primer evolucije na delu je primer glogove muhe Rhagoletis pomonella, za katero se zdi, da je podvržena simpatrični speciaciji.
Različne populacije glogove muhe se hranijo z različnimi plodovi. Posebna populacija se je pojavila v Severni Ameriki v 19. stoletju, nekaj časa po vnosu jabolk, ki so tujerodna vrsta. Ta populacija, ki se hrani z jabolki, se običajno hrani samo z jabolki in ne s plodovi gloga, ki so bili v preteklosti najbolj priljubljeni. Sedanja populacija, ki se hrani z glogom, se običajno ne hrani z jabolki.
Nekateri dokazi kažejo, da prihaja do simpatrične speciacije. Glogove mušice dozorijo pozneje v sezoni in potrebujejo več časa za dozorevanje kot jablanove mušice, poleg tega je malo dokazov o križanju (raziskovalci so zabeležili 4-6-odstotno stopnjo hibridizacije).
Pojav nove glogove muhe je primer napredne evolucije.
Hibridizacija
Redko se zgodi, da nastane nova vrsta, ko se parijo posamezni pripadniki različnih vrst. Običajno so izdelki takšnih križanj neplodni ali relativno neplodni, zato jih naravna selekcija izloči.
Najden je bil en primer nove in uspešne hibridne vrste. Nova vrsta je križanec italijanskega in španskega vrabca, osnovna dejstva pa so bila preverjena z analizo zaporedja DNK v krvi. Ptica živi v Italiji na območju, kjer živita obe starševski vrsti. S španskim vrabcem se ne razmnožuje, čeprav živi z njima drug ob drugem.
Umetna speciacija
Nove vrste so nastale z vzrejo udomačenih živali, vendar začetni datumi in načini niso jasni. Domače ovce so na primer nastale s križanjem in ne dajejo več živahnega potomstva z vrsto Ovis orientalis, iz katere izvirajo.
Po drugi strani pa lahko domače govedo štejemo v isto vrsto kot več vrst divjih volov, gaurjev, jakov itd., saj z njimi zlahka ustvarja plodne potomce.
Lab vrste
Najbolje dokumentirane stvaritve novih vrst v laboratoriju so bile izvedene konec osemdesetih let prejšnjega stoletja. William Rice in G. W. Salt sta vzgojila sadne mušice Drosophila melanogaster z labirintom s tremi različnimi izbirami habitatov, kot so svetlo/temno in mokro/suho.
Vsaka generacija je bila postavljena v labirint, skupine muh, ki so prišle iz dveh od osmih izhodov, pa so bile ločene, da so se razmnoževale med seboj v svojih skupinah. Po petintridesetih generacijah sta bili obe skupini in njuni potomci reproduktivno izolirani zaradi močnih preferenc do habitata: parile so se le na območjih, ki so jim bila ljubša, zato se niso parile z muhami, ki so imele raje druga območja. Zgodovina takšnih poskusov je opisana v Rice in Hostert (1993).
Diane Dodd je lahko tudi pokazala, kako se lahko razmnoževalna izolacija razvije iz preferenc parjenja pri Drosophila pseudoobscura po samo osmih generacijah z uporabo različnih vrst hrane, škroba in maltoze.
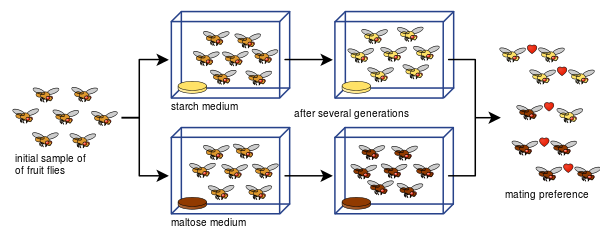
Doddov poskus so zlahka ponovili številni drugi, tudi z drugimi vrstami sadnih mušic in živil.
Okrepitev (učinek Wallace)
Okrepitev je proces, s katerim naravna selekcija povečuje reproduktivno izolacijo.
Pojavi se lahko, ko se dve populaciji iste vrste ločita in nato ponovno prideta v stik. Če je bila njuna reproduktivna izolacija popolna, se bosta že razvili v dve ločeni nezdružljivi vrsti.
Če je njihova razmnoževalna izolacija nepopolna, se bodo pri nadaljnjem parjenju med populacijami pojavili hibridi, ki bodo lahko plodni ali pa tudi ne. Če so hibridi neplodni ali plodni, vendar manj sposobni kot njihovi predniki, potem nadaljnje razmnoževalne izolacije ni več in je do speciacije v bistvu prišlo (npr. pri konjih in oslih).
Razlog za to je, da če imata starša hibridnega potomca naravno izbrane lastnosti za različna okolja, bo hibridni potomec nosil lastnosti obeh in ne bo ustrezal nobeni od niš tako dobro kot oba starša. Nizka primernost hibridov bi povzročila, da bi selekcija dajala prednost asortativnemu parjenju, kar bi zmanjšalo hibridizacijo.
Včasih ga imenujemo Wallaceov učinek po evolucijskem biologu Alfredu Russelu Wallaceu, ki je konec 19. stoletja predlagal, da bi lahko bil pomemben dejavnik pri speciaciji.
Če so hibridni potomci sposobnejši od svojih prednikov, se bosta populaciji na območju, kjer sta v stiku, združili nazaj v isto vrsto.
Za ločevanje je potrebna ojačitev, kadar je med dvema oblikama vrste "hibridno območje". Hibridna območja so območja, kjer se srečujejo in križajo ločene populacije. Na teh območjih so zelo pogosti hibridni potomci, ki običajno nastanejo zaradi sekundarnega stika med divergentnimi vrstami.
Brez okrepitve bi se populaciji ali vrsti nenadzorovano križali. Okrepitev se lahko povzroči v poskusih umetne selekcije, kot je opisano zgoraj.
Vprašanja in odgovori
V: Kaj je speciacija?
O: Specifikacija je proces nastajanja vrst. Je glavni del evolucijske biologije.
V: Kaj je Darwin menil o nastanku vrst?
O: Darwin je menil, da je večina vrst nastala neposredno iz že obstoječih vrst, kar je imenoval anageneza ali "fitotska evolucija".
V: Kako so na splošno gledali na nastanek vrst v 20. stoletju?
O: V 20. stoletju so znanstveniki menili, da je večina vrst nastala, ko so se prejšnje vrste razdelile, kar je znano kot kladogeneza. Menili so tudi, da so to cepitev povzročili ali pomagali povzročiti izolacijski mehanizmi, kot je fizična ločitev.
V: Kako so nedavne raziskave spremenile naše razumevanje speciacije?
O: Nedavne raziskave so pokazale, da lahko pride do križanja med sorodnimi vrstami in da se geni lahko prenašajo s temi križanji. To pomeni, da razmnoževalna izolacija ni vedno potrebna za opredelitev vrste in da alopatrija (fizična ločitev) morda ni potrebna za nastanek vrste.
V: Kaj ima fizična ločitev opraviti z nastankom novih vrst?
O: Fizična ločitev med nekoč soobstoječimi vrstami naj bi bila glavni dejavnik pri nastajanju novih vrst, kar dokazujejo številni primeri, obravnavani v znanstveni literaturi.
Sorodni članki
Avtor
AlegsaOnline.com Speciacija: nastanek vrst v evoluciji, mehanizmi in primeri Leandro Alegsa
URL: https://sl.alegsaonline.com/art/92570
Viri
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833